К вопросу о восприятии времени при зрительно-моторной координации
Кафедра физиологии человека и животных Ростовского госуниверситета
Исследование психофизиологических механизмов перцептивного анализа времен-ных параметров стимулов представляет существенный интерес в понимании сенсомоторной координации. Используемые методические подходы для изучения данных процессов в ос-новном связаны с оценкой длительности различных интервалов времени при помощи срав-нения их с эталонными параметрами. Оценка малых интервалов (менее 1-2 с) представляет собой достаточно простую тестовую процедуру, которая соответствует поставленной задаче. Предъявление длительных интервалов связано с использованием испытуемым метода "внут-реннего отсчета", который представляет собой сложную вербализированную задачу, нейро-физиологический механизм которой в значительной степени маскирует сам процесс воспри-ятия и анализа временных параметров стимуляции. Аналогичные проблемы, но с преобла-данием образного анализа характерны для субъективной оценки заполненности интервала времени набором сенсорных ощущений, длительность которых соответствует измеряемому интервалу.
Использование метода связанных с событием потенциалов (ССП) привело к появ-лению понятия об эндогенных вызванных потенциалах (ВП), возникновение которых опре-деляется не конкретным внешним воздействием, а ожиданием стимула или реакцией в опре-деленный момент времени. К таким ответам относятся ВП на пропущенный стимул, нега-тивная волна ожидания и т.д. [1-4]. В данных тестовых процедурах стимулом для возникно-вения ВП являются интервалы времени, которые измеряются "биологическими часами" сен-сорных систем мозга ("перцептивное время" [5]) и являются основой для прогноза возник-новения реакции ("опережающее отражение" [6]).
Временные параметры стимула (ритмичность, аритмичность, частота, длительность) также являются важными при возникновении экзогенных (вызванных конкретным стиму-лом) ВП [2]. Увеличение длительности межстимульного интервала (МСИ) в диапазоне 1-9 с приводит к увеличению ВР [7].
Таким образом, используя метод ССП, можно говорить о времени как о самостоя-тельном виде стимуляции, и об экзогенных и эндогенных механизмах его отсчета в процессе перцепции, что и явилось предметом настоящего исследования на примере зрительных и слуховых сенсомоторных реакций.
МЕТОДИКА
В исследовании приняло участие 9 человек в возрасте от 20 до 24 лет. В ходе элек-трофизиологического тестирования испытуемые сидели в кресле перед дисплеем в осве-щенной комнате. Расстояние от глаз до экрана составляло примерно 60 см. Стимулом слу-жил белый квадрат со стороной 11,2 мм (40 пикселов), предъявляемый в центре на фоне темного дисплея. Время экспозиции составляло 50 мс. МСИ изменяли от 0.5 до 8 с по лога-рифмической шкале с шагом Ц`2. Всего было использовано 9 интервалов: 500, 707, 1000, 1414, 2000, 2828, 4000, 5687 и 8000 мс. Каждый из указанных МСИ предъявлялся в ходе од-ной серии и был либо периодическим, либо величина его варьировала в пределах + 10%.
ВП регистрировались в отведениях F3, F4, P3, P4, O1 и O2. Референтные электроды располагались на мочках ушей, а индифферентный - на лбу. Комплекс частот около 50 Гц вырезался с помощью режекторного фильтра. Коэффициент подавления синфазного сигнала составлял 105 дБ.
Эпоха анализа составляла 50 мс перед стимулом и до 950 мс после. Перед усредне-нием исходная ЭЭГ фильтровалась с полосой пропускания 0,05-30 Гц. Зрительные ВП ус-реднялись по 50 реализациям. Из суммы исключались ответы с ошибочной ЗМР (за ошибку принимались ответы с ЛП меньше 100 и больше 500 мс), а также те реализации, в процессе которых с помощью электроокулограммы регистрировались значительные векодвигатель-ные реакции.
Испытуемым предлагались следующие тестовые процедуры: (1) Регистрация ВП и/или времени реакции (ВР) при простой зрительно-моторной реакции (ЗМР) при соответ-ствующих длительностях МСИ. (2) При длительностях более 1 с (2, 4, 8 с) испытуемый дол-жен был мысленно производить отсчет времени в секундах, с целью оптимального прогно-зирования момента появления пускового стимула. (3) При подаче ритмической стимуляции частотой 1 Гц испытуемый должен был реагировать в соответствующей серии на 2-й, 4-й или 8-й в соответствии с тестовой процедурой. (4) В ситуации 3 испытуемый должен был осуществлять левой рукой нажатие на каждый стимул, а правой только на 2-й, 4-й или 8-й стимул в соответствии с тестовой процедурой. Аналогичные исследования были проведены для звукового стимула.
Для значений ВР вычисляли среднее, дисперсию и доверительные интервалы. Строились графики зависимости среднего ВР от МСИ, где по оси абсцисс откладывались величина МСИ в логарифмической прогрессии, а по оси ординат - ВР. По этим значениям для каждого испытуемого и при усреднении по исследуемой группе с помощью метода наи-меньших квадратов вычислялись формулы уравнений линейной регрессии y=ax+b (коэффи-циенты a и b).
У ВП оценивались ЛП и амплитуда основных компонентов в диапазоне 150-400 мс. Амплитуда измерялась относительно изолинии. Точность измерения по времени составляла 5 мс, а по амплитуде 0,04 мкВ. Компоненты выделялись визуально.
РЕЗУЛЬТАТЫ ИССЛЕДОВАНИЯ
Использование МСИ от 0.5 до 8 сек показало неоднородность состояний человека при выполнении простой ЗМР, что отражалось в величине дисперсии их ЛП, которая значи-тельно увеличивалась при МСИ меньше 1000 мс с одновременным ростом количества оши-бок (ложная тревога, пропуск стимула). В связи с этим для дальнейшего исследования ис-пользовались МСИ от 1 до 8 с.
У всех испытуемых при увеличении МСИ в выбранном диапазоне было выявлен достоверный рост средней величины ЛП простой ЗМР от 184-320 мс (при 1 с) до 268-410 (при 8 мс) при относительной стабильности дисперсии. Разность между максимальным ЛП и минимальным варьировала в пределах 41-135 мс .
Эта зависимость достоверно апроксимировалась линейным уравнением регрессии (табл. 1). У четырех испытуемых угол наклона прямых примерно одинаков, а свободный член варьировал от 162 до 282 мс. У остальных пяти испытуемых угол наклона прямых меньше, и они имеют приблизительно одинаковую величину свободного члена.
Однородность линейных уравнений регрессии позволила усреднить результаты по всем испытуемым. Увеличение МСИ от 1 до 8 с приводило к достоверному увеличению ЛП простой ЗМР от 241 до 318 мс и имела линейный характер от логарифма длительности МСИ, которую можно описать следующим уравнением:
RT = 0.0272logТ + 0.243
где RT - ЛП простой ЗМР в с, Т - величина МСИ интервала в с.
Из уравнения видно, что увеличение МСИ в 2 раза приводило в среднем к увеличе-нию ЛП на 27 мс, а при МСИ в 1 с составлял 243 мс.
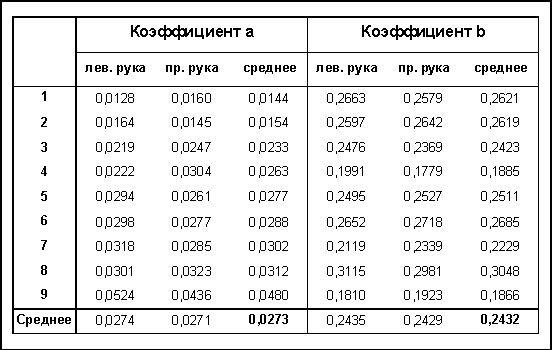
Таблица 1. Индивидуальные коэффициенты регрессии зависимости времени реакции от межстимульного интервала (в секундах)
Для исследования влияния эндогенного отсчета времени на эффективность испы-туемому было предложено с периодичностью в 1 с вести отсчет времени; для экзогенного отсчета - МСИ заполнялся ритмической стимуляцией с частотой 1 Гц. Из рисунка видно, что эндогенный отсчет существенно не повлиял на характер зависимости ЛП простой ЗМР от МСИ. Экзогенный отсчет времени сделал независимым величину ЛП простой ЗМР от вели-чины МСИ.
Одновременный анализ ВП, зарегистрированных на зрительную стимуляцию, яв-ляющейся пусковой для ЗМР, показал, что при увеличении МСИ в исследуемом диапазоне наблюдался рост выраженности ВП (амплитуды и длительности компонентов). Этот процесс регистрировался и в отсутствии двигательной реакции.
Таким образом, увеличение МСИ было связано с увеличением амплитуды ВП и дли-тельности ЛП ЗМР. Аналогичные закономерности были зарегистрированы для акустическо-го воздействия.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Полученные результаты показывают, что увеличение МСИ приводит к увеличению выраженности ВП (особенно поздних компонентов) и ВР. Для ВР эта зависимость носит ло-гарифмический характер. Эти данные согласуются с аналогичными работами [7-9], в кото-рых показано, что существуют короткие и длинные интервалы. Для коротких интервалов характерна линейная прямо пропорциональная зависимость величины ВР от МСИ (интерва-лы до 10 с), для длинных - эта зависимость отсутствует (интервалы более 10 с). Аналогич-ные зависимости получены для параметров ВП, выраженность которых также прямо про-порциональна длительности МСИ [10].
Относительно информационного процесса, лежащего в основе оценки величины МСИ существует несколько точек зрения: (а) запечатление длительности МСИ, заданной внешними стимулами [11,12] (экзогенные "биологические часы"); (б) измерение на основе имеющегося в нервной системе эталона [13] (эндогенные "биологические часы"); (в) изме-рение МСИ осуществляется с использованием обоих механизмов в зависимости от его дли-тельности [14].
Для выяснения преобладающего механизма оценки времени мы использовали две процедуры: (а) устный счет с частотой 1/сек (активация "эндогенных часов"); (б) ритмиче-ский сенсорный стимул с частотой 1/сек (активация "экзогенных часов").
Приведенные выше результаты показали, что при перцептивном анализе времени более эффективными были "экзогенные часы", запускаемые внешними сенсорными (звуко-выми и световыми) стимулами, применение которых существенно уменьшало величину ВР и ВП и делало независимыми их от величины МСИ. Предполагается, что в основе работы "экзогенных часов" лежит динамика уровня активации коры [15], которая может поддержи-ваться двумя возможных механизмами: (а) физиологический (рефрактерность, циклы вос-становления); (б) психологический (ожидание, уровень произвольного внимания).
Для интервалов времени 1-10 с существенным является второй механизм, который при увеличении МСИ приводит к снижению вероятности появления стимула и тем самым к активации механизмов избирательного внимания и, как следствие, к увеличению выражен-ности ВП (особенно поздних компонентов, в частности, компонента Р300). Данные явления характеризуют процесс восприятия.
Для эффекторной реакции наблюдаются противоположные явления, они связаны со снижением активации двигательной системы и увеличением ВР, что можно объяснить рабо-той первого (физиологического) механизма, который отражает взаимоотношения (циклы восстановления) механизмов перцептивной и эффекторной деятельности на коротком ин-тервале времени (200-300 мс). В связи с этим увеличение ВР может быть связано с измене-нием ЛП, амплитуды и длительности компонента Р300, который, по нашим данным [16,17] отражает развитие корковых тормозных процессов, что снижает эффективность двигатель-ной реакции [16].
Таким образом, полученные результаты подтверждают важность временных пара-метров стимулов в процессе зрительно-моторной координации и распознавании образов. На перцептивном уровне при увеличении длительности малых интервалов (1-10 с) преобла-дающими в оценке времени являются "экзогенный часы", которые связаны с процессами активации произвольного внимания, эффективность которого отражается в увеличении ам-плитудно-временных параметров ВП (особенно компонента Р300). При этом наблюдается подавление эффекторной части зрительно-моторной реакции за счет "психологической реф-рактерности" [18], развивающейся после перцептивной деятельности, в основе которой ле-жат тормозные корковые процессы, связанные с развитием компонента Р300.
Полученные результаты позволяют учитывать в конфигурации ВП и ВР процессы, связанные с анализом временных параметров стимулов и их динамику в симметричных об-ластях коры, которая будет определяться величиной предшествующего МСИ и особенно-стями анализа времени, специфичных для каждого полушария.
СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
1. Vaughan H.G. The relationship of brain activity to scalp recordings of event-related potentials// Average evoked potentials. Methods, results, and evaluations. Washington, D.C.: NASA SP-191, 1969. P. 45-94.
2. Рутман Э.М. Вызванные потенциалы в психологии и психофизиологии. М.: Наука. 1979. 216 с.
3. Picton T.W. Human event-related potentials. Handbook of electroencephalography and clinical neurophysiology. Revised Ser. V. 3. Amsterdam: Elsevier, 1988. 556 p.
4. Гнездицкий В.В. Вызванные потенциалы мозга в клинической практике. Таганрог: Изд-во Таганрогского государственного радиотехнического университета, 1997. 252 с.
5. Брагина Н.Н., Доброхотова Т.А. Функциональные асимметрии человека. Москва: Меди-цина. 1988. 240 с.
6. Анохин П.К. Узловые вопросы теории функциональной системы. Москва: Наука. 1980. 196 с.
7. Treisman M. Temporal discrimination and the indifference interval: implication for a model of the internal clock// Psychol. Monogr, 1963. vol. 77. N 13. P. 372-379.
8. Chattergea R.G. Time gap in the estimation of long duration// Percept. and Motor Skills. 1963. Vol. 17 P. 143-149.
9. Chattergea R.G., Sircas A.K. Practice effect in the estimation of short temporal duration// J. Psychol. Res. 1961. Vol. 5. P. 81-84.
10. Кеванишвили З.М., Франганг Б., Хачидзе О.А. и др. Зависимость параметров ранних и поздних колебаний корковых вызванных потенциалов человека от ритма звуковой сти-муляции\\ Журн. высш. нервн. деят. 1976. Т. 26. вып. 1. С. 153.
11. Goldfarb J.E., Goldstone E.S. Proprioceptive involvement, psychophysical method and temporal judgment// Percept. and Motor Skills. 1963. Vol. 17. P. 286.
12. Goldfarb J.E., Goldstone E.S. Time judgment: a comparison of filled and unfilled duration// Percept. and Motor Skills. 1963. Vol. 16. P. 376.
13. Садов Г.А. Оценка коротких временных интервалов по эталонам памяти\\ Новые иссле-дования в психологии. М. 1978. вып. 2. С. 105-109.
14. Алексеев М.А., Крылов Н.В., Лившиц М.П., Найдель А.В. О механизмах координации ритмических движений\\ Вопр. психологии. 1965. № 5. С. 82-89.
15. Суворов Н.Ф., Таиров О.П. Психофизиологические механизмы избирательного внима-ния. Л.: Наука. 1985. 287 с.
16. Айдаркин Е.К. Динамика межанализаторных отношений в процессе выработки оборони-тельного условного рефлекса: Дис. ... канд. биол. наук. М., 1986. 205 с.
17. Айдаркин Е.К. К вопросу об идентификации и природе зрительных вызванных потен-циалов. (Препринт./НИИ нейрокибернетики им. А.Б.Когана Ростовского гос-университета.: N3-96). Изд-во Ростовского университета. Ростов-на-Дону, 1996. 33 с.
18. Welford A.T. Single channel operation in the brain// Acta Psychol. 1967. Vol. 27. P. 23-34.